EzoBox.ruБиблиотека эзотерики
В 1960 г. нами (Уголев, 1960б) было продемонстрировано, что экстракты двенадцатиперстной кишки, но не желудка, печени, мышц и селезенки, вызывают специфическое торможение аппетита (рис. 8.1). После введения экстракта двенадцатиперстной кишки поведение голодных крыс напоминало поведение сытых животных. Снижение потребления пищи наблюдалось после введения таких экстрактов как сытым, так и голодным животным. На основании этих данных можно допустить, что существует кишечный гормон или фактор, осуществляющий торможение аппетита, т.е. торможение той важнейшей реакции, которая характеризует изменение поведения человека и животных после удовлетворения пищевых потребностей. В 1962 г. (Уголев, 1962) были получены сведения, которые позволили предположить, что такой фактор, скорее всего, является пептидом. Наличие дуоденального фактора, вызывающего сытость, который был назван нами арэнтерином (аппетитрегулирующим энтерином, т.е. кишечным гормоном), так же как и его пептидная природа получили подтверждение во многих работах. Все же следует отметить, что до сих пор арэнтерин в чистом виде не получен.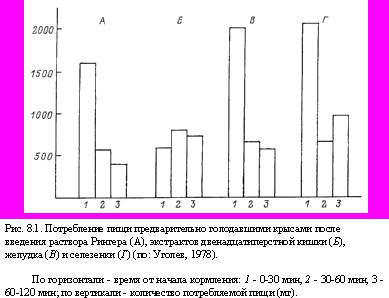
![]()
Попытки охарактеризовать арэнтерин были предприняты группой чл.-корр. АН СССР А. С. Хохлова в Институте биоорганической химии им. М. М. Шемякина АН СССР и в нашей лаборатории. В середине 70-х годов обеими группами было продемонстрировано, что арэнтерин - это белок с молекулярной массой около 100000 (обзоры: Уголев, 1978, 1985).
8.4.2. Другие кишечные гормоны
В 70-х годах было обнаружено, что многие известные гормоны желудочно-кишечного тракта, в частности гастрин и секретин, не оказывают влияния на аппетит. В отличие от них холецистокинин вызывает сильное торможение пищевой активности. Первым, по-видимому, продемонстрировал специфическое влияние холецистокинина на потребление пищи X. Купманс с сотрудниками (Koopmans et al., 1972). В цикле исследований группы Дж. Смита (Smith et al., 1974) на крысах было показано, что холецистокинин может вызывать торможение аппетита. Авторы, однако, оценивали этот эффект, скорее, как фармакологический, чем физиологический. у обезьян также обнаружено торможение потребления пищи после введения холецистокинина. У человека потребление пищи зависит от дозы введенного холецистокинина и от способа его введения. Так, внутривенное введение гормона в дозе 0.5 ед/кг приводит к торможению аппетита, а постепенное внутривенное введение в дозе 1 ед/кг, напротив, вызывает его стимуляцию (Sturdevant, Goetze, 1976). Прямое действие холецистокинина на центры питания и другие нервные центры показано в электрофизиологическом исследовании Н. Дафни и Е. Джекобсона (Dafny, Jacobson, 1975). Наконец, продемонстрировано, что после введения глюкагона возрастает электрическая активность центра сытости. (Более подробно эти вопросы освещены в монографии: Уголев, 1978.).
В 80-е годы интенсивная работа в этом направлении продолжалась. На человеке и животных было вновь продемонстрировано, что в передаче сигнала насыщения большую роль играют кишечные гормоны. Суммируя полученные за эти годы данные, можно сказать, что экзогенный холецистокинин, в том числе его синтетический октапептид холецистокинин-8, вызывает снижение потребления пищи, которое зависит от дозы гормона и места его введения (периферического или центрального). При этом для развития эффекта насыщения необходим сульфатированный тирозин в 7-й позиции С-терминального конца цепи октапептида холецистокинина. Показана также физиологическая специфичность холецистокинина: он ингибирует потребление пищи, но не воды (Smith, 1983; Gibbs, Smith, 1986, и др.).
В последние годы дискутируется вопрос о периферическом и центральном механизмах действия холецистокинина на аппетит. (Физиологический и фармакологический эффекты гормона дифференцировать достаточно трудно.)
Так как систематические инъекции холецистокинина приводят к резкому изменению пищевого поведения, то предполагается, что гормон должен прямо или опосредованно вызывать изменения функционального состояния мозговых структур. Считается, что основным путем передачи информации после введения экзогенного холецистокинина служат афферентные волокна абдоминальной вагусной системы, с рецепторами которой связывается гормон. Кроме того, гормон может связываться и с рецепторами пилорического сфинктера желудка.